
 Quanto à questão das origens humanas, as posições de vários escritores estão relacionadas a sua colocação em uma das quatro categorias gerais que definem o assunto mais amplo das origens (Schaefer, Ciência e Cristianismo: Conflito ou Coerência, 2003):
Quanto à questão das origens humanas, as posições de vários escritores estão relacionadas a sua colocação em uma das quatro categorias gerais que definem o assunto mais amplo das origens (Schaefer, Ciência e Cristianismo: Conflito ou Coerência, 2003):
1) Evolução naturalista.
Processos impessoais, por exemplo, seleção natural, mutações, acaso, ou alguma combinação destes, são responsáveis por todas as formas e espécies de vida. Se emerge gradualmente (Dawkins) ou aparece de repente (Gould), a humanidade é produto de forças impensadas e não-intencionais.
2) Evolução teísta.
Deus como agente imanente sustenta e direciona os processos naturais que moldam a evolução da vida. Esta posição é cientificamente indistinguível da primeira, mas pressupõe a atividade soberana de Deus em planejar e executar o processo evolutivo.
3) Criação progressiva.
Deus imanentemente direciona um desenvolvimento extensivo das espécies. Deus age transcendentalmente nos estágios especiais deste processo para criar as principais ordens biológicas do ser. A humanidade não é dependente fisicamente de nenhuma espécie intermediária. A idade do universo é cerca de 13,8 bilhões de anos, e a idade da Terra é cerca de 4,7 bilhões de anos.
4) Criação recente.
Todas as formas de vida são criadas “de novo” (do começo) pela Agência sobrenatural. Nenhuma ordem tardia da criação é dependente dos tipos anteriores de seres. A idade da Terra não é mais que 10 mil anos.
Fonte: http://www.asa3.org/ASA/topics/Book%20Reviews2005-/12-05.html
Obviamente, “evolução naturalista” é incompatível com a fé cristã. Cada vez mais, cristãos informados cientificamente estão reconhecendo que a “criação recente” é incompatível com a evidência científica e requer a assunção de “criação com uma aparência de idade”. Tal perspectiva é frequentemente vista como filosoficamente insatisfatória porque isso implica engano na parte do Criador. Conformemente, as únicas perspectivas viáveis para muitos apologistas cristãos são a “evolução teísta” ou a “criação progressiva”. Muitos são hesitantes em considerar a “evolução teísta” porque é “cientificamente indistiguível” da “evolução naturalista”. Do mesmo modo, é frequentemente considerado exigir compromisso de doutrinas bíblicas amplamente difundidas a respeito da criação. Por estas razões, a “criação progressiva” é cada vez mais vista como a perspectiva mais convincente entre os apologistas cristãos modernos. A evidência da popularidade desta perspectiva é refletida no crescente interesse pelo “Design Inteligente” (DI) bem como a popularidade de várias publicações apologéticas tais como aquelas publicadas pelo “The Discovery Institute” e “Reasons to Believe”.
A recente realização do “Projeto Genoma Humano” em 2003 providenciou acesso à informação que é relevante para a questão das origens humanas e apologética cristã. O diretor do projeto, Francis Collins, abandonou o ateísmo e abraçou a fé cristã em 1977. Ele é o autor de “A linguagem de Deus”, que foi publicado em julho de 2006. Nesse sentido, Dr. Collins está em uma posição única para comentar sobre as questões de ciência e fé quando são relacionadas à questão das origens humanas. A informação que se segue merece consideração por apologistas cristãos que estão interessados na controvérsia associada aos tópicos de criação e evolução. As páginas seguintes contêm extratos de “A linguagem de Deus” que são considerados relevantes para essa questão. Certas seções são destacadas em negrito para realçar alguns dos pontos relevantes feitos por Collins no que se refere à questão das origens humanas.
“Livros inteiros foram escritos sobre o Projeto Genoma Humano (provavelmente muitos, de fato). Talvez eu escreva o meu próprio algum dia, esperançosamente com retrospectiva suficiente para evitar alguns dos pronunciamentos esbaforidos de muitas das atuais representações populares. Não é o propósito deste livro, no entanto, alongar-se mais nessa experiência notável, mas refletir sobre os caminhos nos quais uma compreensão moderna da ciência pode ser harmonizada com uma crença em Deus.
A esse respeito, é interessante olhar com cuidado o genoma da humanidade, e compará-lo com os genomas de muitos outros organismos que foram agora sequenciados. Quando nós pesquisamos a vasta extensão do genoma humano, 3,1 bilhão de letras do código de DNA arranjam-se sobre vinte e quatro cromossomos, muitas surpresas são imediatamente aparentes.
Outra característica marcante do genoma humano vem da comparação de diferentes membros da nossa própria espécie. Ao nível do DNA, somos todos 99,9% idênticos. Essa semelhança aplica-se independentemente de quais dois indivíduos de todo o mundo você escolhe comparar. Assim, pela análise de DNA, nós humanos somos verdadeiramente parte de uma família. Esta notável baixa diversidade genética nos distingue da maioria das outras espécies no planeta, onde a quantidade de diversidade do DNA é dez ou, às vezes, até cinquenta vezes maior que a nossa.
Geneticistas populacionais, cuja disciplina envolve o uso de ferramentas matemáticas para reconstruir a história das populações de animais, plantas, ou bactérias, olham para estes fatos sobre o genoma humano e concluem que eles apontam que todos os membros da nossa espécie descenderam de um conjunto comum de fundadores, aproximadamente 10 mil em número, que viveram cerca de 100 mil a 150 mil anos atrás. Esta informação se adequa bem com o registro fóssil, que por sua vez coloca o local daqueles ancestrais fundadores provavelmente na África Oriental.
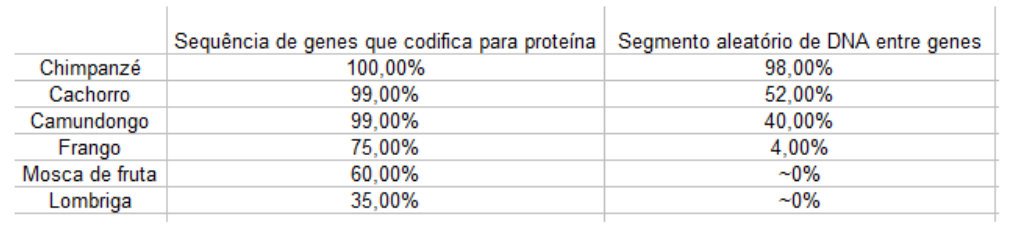
Outra consequência profundamente interessante do estudo de genomas múltiplos foi a habilidade de fazer comparações detalhadas de nossa própria sequência de DNA com a de outros organismos. Usando um computador, pode-se pegar um certo trecho do DNA humano e avaliar se há uma sequência semelhante em alguma outra espécie. Se alguém pega a região codificadora do gene humano (isto é, a parte que contém as instruções para uma proteína), e usa-a para a pesquisa, haverá quase sempre uma correspondência altamente significativa aos genomas de outros mamíferos. Muitos genes se mostrarão discerníveis, mas imperfeitos correspondentes aos peixes. Alguns até encontrarão correspondentes aos genomas de organismos mais simples, tais como moscas de frutas e parasitas intestinais. Em alguns exemplos particularmente marcantes, a semelhança se estenderá por todo o caminho aos genes em levedura e até às bactérias.
Se, por outro lado, alguém escolhe um pedaço do DNA humano que se encontra entre genes, então a possibilidade de encontrar uma sequência semelhante nos genomas de organismos distantemente relacionados diminui. Não desaparece inteiramente; com uma pesquisa cuidadosa de computador, cerca de metade de tais fragmentos pode ser alinhada com outros genomas de mamíferos, e quase todos eles alinham-se bem com o DNA de outros primatas não-humanos. A tabela 5.1¹ mostra as porcentagens do sucesso deste tipo de combinação, dividida em várias categorias.
O que tudo isto significa? Em dois níveis diferentes, fornece poderoso apoio para a teoria da evolução de Darwin, isto é, descendência de um ancestral comum² com seleção natural operando em variações aleatórias. Ao nível do genoma como um todo, um computador pode construir uma árvore da vida baseado unicamente nas semelhanças das sequências de DNA de múltiplos organismos. O resultado é mostrado na Figura 5.1.
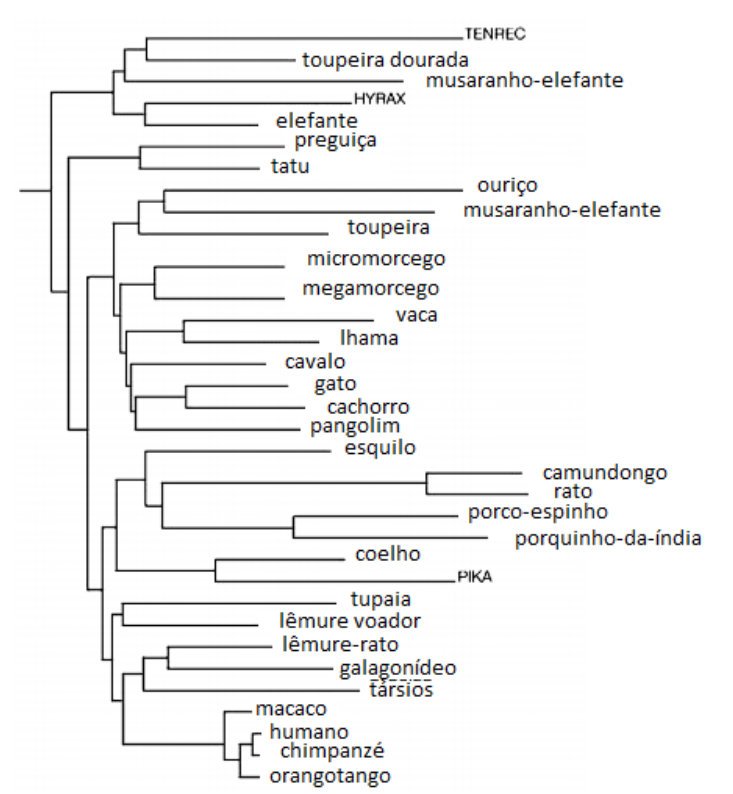
Tenha em mente que esta página não utiliza nenhuma informação dos registros fósseis, ou de observações anatômicas de formas de vida atuais. Ainda esta semelhança às conclusões tiradas de estudos de anatomia comparativa, ambos de organismos existentes e de restos fossilizados, é marcante. Segundo: dentro do genoma, a teoria de Darwin prediz que mutações que não afetam função (a saber, aquelas localizadas no “DNA-lixo”) vão se acumular de forma constante ao longo do tempo. Mutações na região codificadora dos genes, no entanto, deverão ser observadas com menos frequência, uma vez que a maioria será deletéria, e apenas um raro evento vai providenciar uma vantagem seletiva e ser retido durante o processo evolutivo. Isso é exatamente o que é observado. Este último fenômeno até aplica-se aos finos detalhes das regiões codificadoras de genes. Do capítulo anterior, você pode recordar-se de que o código genético é degenerado: por exemplo, GAA e GAG, ambos códigos para ácido glutâmico. Isso significa que é possível para algumas mutações na região codificadora estarem “silenciosas”, onde o aminoácido codificado não é alterado pela mudança, e então nenhuma pena é paga. Quando compara-se sequências de DNA de espécies relacionadas, diferenças silenciosas são muito mais comuns nas regiões codificadoras do que aquelas que alteram um aminoácido. É exatamente o que a teoria de Darwin prediria. Se, como alguns podem argumentar, estes genomas foram criados por atos individuais de criação especial, porque esta característica particular apareceria?
Charles Darwin eram intensamente inseguro sobre sua teoria da evolução. Talvez isso seja responsável pelos quase vinte e cinco anos que se passaram entre o desenvolvimento da ideia e a publicação de “A origem das espécies”. Deve ter havido muitas vezes nas quais Darwin desejasse poder voltar milhões de anos no tempo e realmente observar todos os eventos que sua teoria previu. Claro que ele não poderia fazer isso, e nós também não podemos fazer hoje. Mas faltando uma máquina do tempo, Darwin dificilmente poderia ter imaginado uma demonstração digital mais convincente de sua teoria do que a que nós encontramos estudando o DNA de organismos múltiplos.
Em meados do século XIX, Darwin não tinha como saber o que seria o mecanismo de evolução por seleção natural. Nós agora podemos ver que a variação que ele postulou é apoiada por mutações naturais no DNA. Estas são estimadas a ocorrer a uma taxa de cerca de um erro a cada 100 milhões de pares de bases por geração. (Isso significa, a propósito, que desde que todos temos dois genomas com 3 bilhões de pares de base cada, um de nossa mãe e um de nosso pai, todos temos cerca de sessenta novas mutações que não foram presentes em nenhum dos nossos pais.)
A maioria daquelas mutações ocorre em partes do genoma que não são essenciais, e portanto elas têm pequena ou nenhuma consequência. Aquelas que caem nas partes mais vulneráveis são geralmente nocivas, e são assim rapidamente abatidas da população porque elas reduzem a capacidade reprodutiva. Mas em raras ocasiões, uma mutação vai surgir por acaso que ofereça um leve grau de vantagem seletiva. Essa nova “ortografia” do DNA terá uma possibilidade levemente maior de ser passada para futura descendência. Ao longo de um período muito longo de tempo, tais raros eventos favoráveis podem se tornar muito difundidos em todos os membros da espécie, por fim resultando em grandes alterações na função biológica.
Em algumas instâncias, cientistas estão até capturando a evolução no flagrante, agora que temos as ferramentas para rastrear estes eventos. Alguns críticos do Darwinismo gostam de argumentar que não há evidência da “macroevolução” (isto é, grande alteração na espécie) no registro fóssil, apenas de “microevolução” (mudança incremental dentro de uma espécie). Temos visto bicos de tentilhões mudarem de forma ao longo do tempo, eles argumentam, dependendo da mudança das fontes de alimento, mas não temos visto novas espécies surgirem. Esta distinção é cada vez mais vista como artificial.
Desde o tempo de Darwin, pessoas de muitas cosmovisões diferentes foram particularmente motivadas a entender como revelações sobre biologia e evolução se aplicam a essa classe especial de animais, os seres humanos. O estudo dos genomas leva inexoravelmente à conclusão de que nós humanos partilhamos um ancestral comum com outros seres vivos. Algumas dessas evidências são mostradas na Tabela 5.1, onde a semelhança entre nossos genomas e os de outros organismos é exibida. Esta evidência sozinha, é claro, não prova um ancestral comum; da perspectiva de um criacionista, tais semelhanças poderiam simplesmente demonstrar que Deus usou princípios de design de sucesso repetidas vezes. Como devemos ver, no entanto, e como foi prefigurado acima pela discussão das mutações “silenciosas” em regiões codificadoras de proteína, o estudo detalhado dos genomas tornou essa interpretação praticamente insustentável – não só sobre todos os seres vivos, mas também sobre nós mesmos.
Como um primeiro exemplo, vamos olhar para uma comparação dos genomas do humano e do camundongo, ambos os quais foram determinados à alta precisão. O tamanho total dos dois genomas é grosseiramente o mesmo, e o inventário de genes codificadores de proteína é notavelmente parecido. Mas outros sinais inequívocos de um ancestral comum rapidamente aparecem quando alguém olha os detalhes. Por exemplo, a ordem dos genes ao longo dos cromossomos humanos e do rato é geralmente mantida ao longo de extensões consideráveis de DNA. Assim, se encontro genes humanos A, B e C nessa ordem, é provável descobrir que os do camundongo têm homólogos de A, B e C também colocados nessa mesma ordem, apesar de o espaçamento entre os genes ter variado um pouco (Figura 5.2).
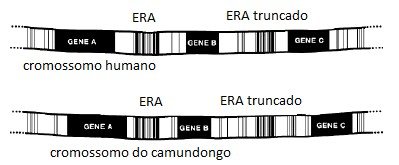
Uma evidência até mais convincente de um ancestral comum vem do estudo do que são conhecidos como elementos repetitivos antigos (ERAs). Estes surgem dos “genes saltadores”, que são capazes de copiar e inserir-se em diversos outros locais no genoma, geralmente sem consequências funcionais. Genomas de mamíferos estão repletos de tais ERAs, com cerca de 45% do genoma humano feito destes destroços genéticos. Quando alguém alinha seções dos genomas do homem e do camundongo, ancorado pelo aparecimento de homólogos de genes que ocorrem na mesma ordem, pode-se geralmente também identificar ERAs aproximadamente na mesma localização nestes dois genomas (Figura 5.2).
Alguns destes podem ter se perdido em uma espécie ou outra, mas muitos deles permanecem em uma posição que é mais consistente com o ter chegado no genoma de um ancestral comum de mamíferos, e terem sido levados desde então. Claro, alguns podem argumentar que estes são na verdade elementos funcionais colocados lá pelo Criador por uma boa razão, e que nosso desconto deles como “DNA-lixo” apenas trai nosso atual nível de ignorância. E de fato, uma pequena fração deles pode desempenhar importantes papéis reguladores. Mas certos exemplos severamente esforçam a credulidade dessa explicação. O processo de transposição frequentemente danifica o gene saltador. Há ERAs ao longo dos genomas do homem e do camundongo que foram truncados quando chegaram, removendo qualquer possibilidade de funcionamento. Em muitos casos, pode-se identificar um ERA decapitado e totalmente extinto em posições paralelas no genoma humano e do camundongo (Figura 5.2).
A menos que se esteja disposto a assumir a posição de que Deus colocou esses ERAs decapitados nestes lugares precisos para confundir-nos e enganar-nos, a conclusão de um ancestral comum para humanos e camundongos é praticamente inevitável. Este tipo de dados recentes do genoma assim apresenta um desafio esmagador para aqueles que mantêm a ideia de que todas as espécies foram criadas ex nihilo.
O lugar dos humanos na árvore da vida evolutiva é apenas reforçado por uma comparação com nosso parente vivo mais próximo, o chimpanzé. A sequência do genoma do chimpanzé agora foi revelada, e revela que humanos e chimpanzés são 96% idênticos no nível do DNA.
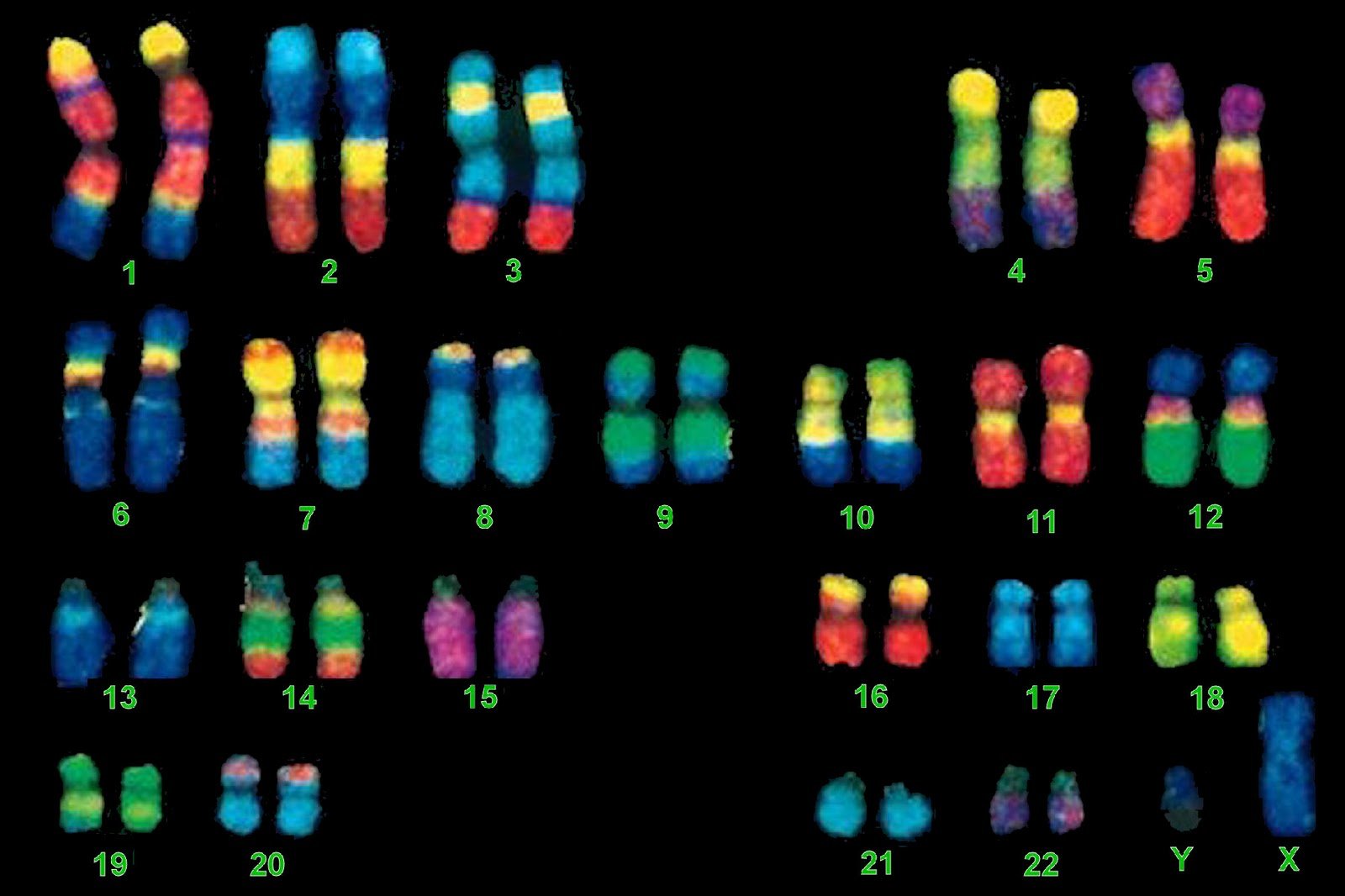
Outro exemplo desta relação próxima se origina do exame da anatomia dos cromossomos humanos e do chimpanzé. Cromossomos são a manifestação visível do genoma do DNA, aparentes no microscópio de luz no momento em que a célula se divide. Cada cromossomo contém centenas de genes. A figura 5.3 mostra uma comparação dos cromossomos entre um humano e um chimpanzé.
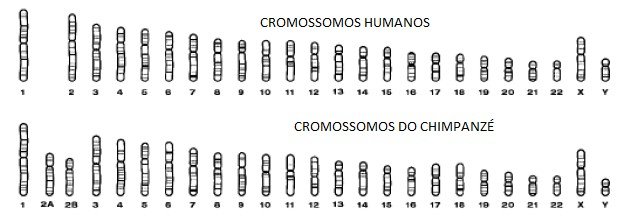
O homem tem 23 pares de cromossomos, mas o chimpanzé tem 24. A diferença no número de cromossomos parece ser a consequência de dois cromossomos ancestrais fundidos para gerar o cromossomo humano 2. Que o do homem pode ser uma fusão é sugerido ao estudar o gorila e orangotango – cada um tem 24 pares de cromossomos, muito parecidos com o chimpanzé.
Recentemente, com a determinação da sequência completa do genoma humano, tornou-se possível olhar para a localização precisa onde esta proposta fusão cromossômica deve ter acontecido. A sequência neste local – ao longo do braço longo do cromossomo 2 – é verdadeiramente notável. Sem entrar nos detalhes técnicos, deixe-me dizer que as sequências especiais ocorrem nas pontas dos cromossomos de todos os primatas. Essas sequências geralmente não ocorrem em outro lugar. Mas são encontradas exatamente onde a evolução previria, no meio do nosso segundo cromossomo fundido. A fusão que ocorreu quando evoluímos dos macacos deixou sua marca de DNA aqui. É muito difícil entender esta observação sem postular um ancestral comum.
Ainda outro argumento para a ascendência comum de chimpanzés e humanos vem da observação peculiar do que são chamados pseudogenes. Esses são genes que têm quase todas as propriedades de um pacote de instruções do DNA funcional, mas são atingidos por uma ou mais falhas que transformam seu script em linguagem sem nexo. Quando compara-se o chimpanzé e o humano, genes ocasionais parecem ser claramente funcionais em uma espécie mas não em outra, porque adquiriram uma ou mais mutações deletérias. O gene humano conhecido como Caspase-12, por exemplo, sustentou vários golpes de nocaute, apesar de ser encontrado na localização relativa idêntica no chimpanzé. O gene Caspase-12 do chimpanzé funciona bem, assim como o gene semelhante em quase todos os mamíferos, inclusive os camundongos. Se os humanos surgiram como uma consequência de um ato sobrenatural de criação especial, porque Deus se daria ao trabalho de inserir tal gene não-funcional neste local preciso?
Nós podemos também agora começar a explicar as origens de uma pequena fração das diferenças mais mecânicas entre nós e nossos parentes mais próximos, algumas delas que podem desempenhar papéis cruciais em nossa humanidade. Em um exemplo, um gene para a proteína do músculo da mandíbula (MYH 16) parece ter sofrido uma mutação para um pseudogene nos humanos. Continua a desempenhar um papel importante no desenvolvimento e força dos músculos da mandíbula em outros primatas. É concebível que a inatividade deste gene levou a uma redução na massa do músculo da mandíbula humana. A maioria dos macacos têm relativamente mandíbulas maiores e mais fortes do que as nossas. Os crânios do homem e do macaco devem, entre outras coisas, servir como uma âncora para estes músculos da mandíbula. É possível que o desenvolvimento de mandíbulas mais fracas paradoxalmente permitiu que nossos crânios se expandissem para cima, e acomodassem nossos cérebros maiores. Isto é claramente especulação, claro, e outras mudanças genéticas seriam necessárias para ter em conta o córtex cerebral muito maior que representa um componente principal da diferença entre humanos e chimpanzés.
Em outro exemplo, muito interesse tem recentemente rodeado o gene chamado FOXP2 por causa de seu papel potencial no desenvolvimento da linguagem. A história do FOXP2 começou com a identificação de uma única família na Inglaterra onde os membros de três gerações tinham grave dificuldade na fala. Eles lutavam para pronunciar palavras de acordo com as regras gramaticais, para entender estrutura complexa de frases, e mover os músculos de suas bocas, rostos, e caixas de voz, para articular certos sons.
Em um grande esforço de investigação genética, os membros afetados da família descobriram ter uma única letra do código de DNA com erros no gene FOXP2 no cromossomo 7. O fato de que um único gene com um sutil erro de ortografia poderia causar tais déficits de linguagem profundos, sem outras consequências óbvias, foi totalmente surpreendente.
A surpresa rapidamente aumentou quando foi mostrado que a sequência deste mesmo gene FOXP2 manteve-se notavelmente estável em quase todos os mamíferos. A mais dramática exceção, no entanto, são os humanos onde duas alterações significativas ocorreram na região codificadora do gene, aparentemente tão recentemente quanto cem mil anos atrás. A hipótese sugerida por estes dados é que estas mudanças recentes no FOXP2 podem de alguma maneira ter contribuído para o desenvolvimento da linguagem nos seres humanos.
Neste ponto, materialistas ateus devem estar animados. Se os humanos evoluíram estritamente por mutação e seleção natural, quem precisa de Deus para explicar-nos? A isto, eu respondo: eu preciso. A comparação das sequências do chimpanzé e do humano, interessante como é, não nos diz o que significa ser humano. Na minha opinião, a sequência de DNA sozinha, mesmo se acompanhada por um vasto tesouro de dados sobre a função biológica, nunca explicará certos atributos humanos especiais; tais como o conhecimento da Lei Moral e a busca universal por Deus. Libertar Deus do fardo de atos especiais de criação não O remove como fonte das coisas que fazem a humanidade especial, e do próprio universo. Simplesmente nos mostra algo de como Ele opera.
Estes exemplos apresentados aqui do estudo dos genomas, além de outros que poderiam encher centenas de livros deste comprimento, fornecem o tipo de apoio molecular para a teoria da evolução que convenceu praticamente todos os biólogos que trabalham de que o quadro de variação e seleção natural de Darwin é inquestionavelmente correto. De fato, para aqueles como eu mesmo trabalhando em genética, é quase impossível imaginar correlacionar as vastas quantidades de dados surgindo dos estudos de genomas sem os fundamentos da teoria de Darwin. Como Theodosius Dobzhansky, um biólogo líder do século XX (e um devoto cristão ortodoxo oriental), disse, “nada na biologia faz sentido exceto à luz da evolução”.
Claramente, no entanto, a evolução tem sido a fonte de grande desconforto na comunidade religiosa ao longo dos últimos 150 anos, e essa resistência não mostra sinais de diminuição. Ainda os crentes seriam bem advertidos a olhar cuidadosamente para o peso esmagador dos dados científicos apoiando esta visão do parentesco de todas as coisas vivas, incluindo nós mesmos. Dada a força da evidência, é desconcertante que tão pouco progresso na aceitação do público ocorreu nos Estados Unidos.”³
Em síntese, algumas das conclusões centrais que Collins apresentou no livros são as seguintes:
- Todos os membros da nossa espécie descenderam de um conjunto comum de fundadores, aproximadamente 10 mil em número, que viveram cerca de 100 mil a 150 mil anos atrás.
- O estudo dos genomas leva inexoravelmente à conclusão de que nós humanos partilhamos um ancestral comum com outras coisas vivas.
- Enquanto alguns criacionistas argumentaram que semelhanças nos genomas entre organismos diferentes poderiam simplesmente demonstrar que Deus usou princípios de design de sucesso repetidas vezes, a evidência de mutações “silenciosas” em regiões codificadoras de proteína, juntamente com o estudo detalhado dos genomas, tornou essa interpretação praticamente insustentável – não só sobre todas as coisas vivas, mas também sobre nós mesmos.
- Uma evidência até mais convincente de um ancestral comum vem do estudo do que são conhecidos como elementos repetitivos antigos (ERAs).
- A menos que se esteja disposto a assumir a posição de que Deus colocou ERAs decapitados em posições precisas para nos confundir e nos enganar, a conclusão de um ancestral comum para humanos e outros organismos é praticamente inevitável. Este tipo de dados recentes do genoma apresenta assim um desafio esmagador àqueles que mantêm a ideia de que todas as espécies foram criadas ex nihilo.
- Os exemplos apresentados do estudo de genomas, além de outros que poderiam encher centenas de livros, fornecem o tipo de apoio molecular para a teoria da evolução que convenceu praticamente todos os biólogos que trabalham de que o quadro de variação e seleção natural de Darwin é inquestionavelmente correto.
Claro que estas conclusões são incompatíveis com a perspectiva de Criação Progressiva explicada no livro recente, “Quem foi Adão?”.* Henry F. Schaefer III, professor Graham Perdue de Química, da Universidade da Georgia, creditou o livro como sendo “a contribuição mais importante ao debate das origens humanas a aparecer durante os últimos cinquenta anos”.** (Nota-se que esta crítica foi publicada em setembro de 2005, antes da publicação de A Linguagem de Deus em julho de 2006.) Curiosamente, dr. Fazale (Fuz) Rana, dr. Dave Rogstad, e Joe Aguirre, todos membros do quadro de funcionários do Reasons to Believe, realizaram uma entrevista por telefone com Francis Collins em seu programa de rádio semanal “Creation Update” (n.339) que foi ao ar em 17 de outubro de 2006. (Clique no link abaixo para acessar o arquivo de áudio).
http://c450903.r3.cf2.rackcdn.com/2006/cu339.mp3
Durante a entrevista, dr. Collins fez os seguintes comentários em resposta às várias questões que ele recebeu:
- “Certamente, quando olho para o genoma dos humanos hoje, todos os seis ou sete bilhões de pessoas no planeta, não é consistente ter descendido de um único par de indivíduos. É consistente descender de um grupo maior de talvez 10 mil ancestrais ou mais.”
- Em resposta a uma questão do dr. Rana a respeito da natureza “provocativa” da Teoria Fora da África, em vista do rastreamento genético da origem da humanidade de volta à sequência de um único gene que é referida como Eva mitocondrial e Adão cromossomial-Y, dr. Collins respondeu: “Todo o assunto de Eva mitocondrial e Adão cromossomial-Y, eu penso que tem sido super-ínterpretado por aqueles que não olharam profundamente como a genética da população funciona…A mitocôndria é, afinal, uma parte pequena da quantidade total de DNA. Quando você olha para o resto do DNA, indo além da mitocôndria e do cromossomo Y, está claro que não pode explicar a humanidade moderna com base em dois únicos antepassados, um macho e uma fêmea – não vai funcionar.”
- Rogstad reconheceu que não foi provado que a origem da humanidade pode ser rastreada de volta à sequência de um único gene, mas pressionou dr. Collins se tal conclusão é ou não excluída pela evidência genética ou biológica. Dr. Collins respondeu: “Eu penso que é, na verdade…Não há jeito de chegar aonde estamos hoje a partir daquele conjunto original de quatro cópias (baseado em dois indivíduos). Realmente exige que você postule que havia um grupo maior do que apenas dois. Novamente, não estou certo de que queria promover isto como a resposta que quero, mas é a resposta que vejo dos dados.”
Uma apresentação mais detalhada das conclusões do dr. Collins do Projeto Genoma Humano é fornecida em um vídeo disponível para download do seguinte link:
http://www.asa3.org/ASA/multimedia.html
Enquanto o livro “Quem foi Adão?” é sem dúvida o recurso definitivo que representa a visão da Criação Progressiva, suas críticas da evidência do DNA para a descendência comum geralmente consistem naquelas que o dr. Collins mencionou em seu livro, tais como a funcionalidade real do “DNA-lixo”, etc. Baseado nesta conclusão, dr. Rana argumentou que a localização comparável do “DNA-lixo” dentro dos genomas de diferentes organismos é proposital e não uma evidência de descendência comum. A seguinte citação resume sua análise racional:
“A geografia comum das sequências de DNA não-codificadoras nos genomas humano e do chimpanzé provavelmente se originam do seu papel de regular a atividade do gene. Estas sequências de DNA devem ser posicionadas precisamente (em relação aos genes que elas controlam) para exercer influência adequada.
E sobre o material genético sem uma função conhecida, como os pseudogenes unitários GLO que humanos e chimpanzés compartilham? Atualmente o modelo RTB não oferece explicação para este aspecto. O modelo prevê, no entanto, que como com outras classes de DNA não-codificador, a função um dia será descoberta para estes pseudogenes de união.”***
Mais tarde no programa, dr. Rana defendeu sua posição para um interlocutor, fazendo diversas referências a “Quem foi Adão?”.
Como mencionado anteriormente, dr. Collins não considera que o argumento RTB explique satisfatoriamente os ERAs “decapitados” e “completamente extintos” em posições paralelas nos genomas de humanos e outros organismos. Em sua perspectiva, tal argumento implica uma ação enganosa da parte do Criador não muito diferente dos “raios de luz criados no trânsito” que são associados com a posição criacionista da Terra jovem.
Em vista da evidência convincente do Projeto Genoma Humano apoiando a evolução (incluindo a descendência comum), apologistas cristãos modernos deveriam considerar as implicações. Embora há certamente muitos defensores da evolução que são ateístas ou agnósticos, há também muitos defensores contemporâneos do criacionismo evolucionista que também abraçaram a fé cristã. Dois livros relativamente recentes representando a perspectiva de tais crentes incluem “Finding Darwin’s God” (Encontrando o Deus de Darwin) e “Perspectives on an Evolving Creation” (Perspectivas sobre uma criação em evolução). A evolução não é mais uma teoria que pode simplesmente ser relegada a uma conspiração de incrédulos. Além disso, existe a erudição bíblica legítima que fornece muitas respostas relevantes para a conciliação da revelação geral (natureza) e revelação especial (escritura). Um exemplo de um livro que é útil a esse respeito é “Inspiration and Incarnation” (Inspiração e encarnação).
Hoje, ainda há muitos crentes que concordam com o criacionismo da Terra jovem. De maneira semelhante, ainda há crentes que consideram a versão do Rei Jaime (King James Version) da Bíblia como a única interpretação confiável da Palavra de Deus. A porcentagem de defensores de “apenas KJV” certamente diminuiu à medida que crentes em geral tornaram-se mais educados sobre o processo de tradução e a superioridade real (em vez de inferioridade) de muitas traduções modernas (comparadas à exatidão da KJV). Do mesmo modo, quanto mais crentes tornaram-se mais instruídos cientificamente e biblicamente, eles perceberam que a aceitação de uma perspectiva de Terra jovem não é essencial para manter uma fé cristã ortodoxa. Ademais, muitos perceberam que a revelação geral de Deus claramente aponta para uma Terra que tem bilhões (não milhares) de anos.
Aqueles que insistem que a KJV é a única tradução legítima da Bíblia, juntamente com aqueles que insistem que a admissão de uma Terra jovem é essencial à fé, ambos colocam uma “pedra de tropeço” no caminho dos incrédulos (os últimos provavelmente mais do que os primeiros). Como embaixadores de Cristo, temos a obrigação de ajudar as pessoas a desenvolver a fé – não impedir através da imposição de obstáculos ilegítimos. Parece que a maioria aprendeu esta lição no que se refere à questão da KJV. Cada vez mais, muitos estão aprendendo esta lição no que se refere à idade da Terra. É o momento para os apologistas cristãos modernos também aprenderem esta lição no que se refere à questão da evolução. Embora esta questão atualmente pode ter aplicação limitada devido à falta geral de consciência sobre as implicações do Projeto Genoma Humano, este não será sempre o caso. À medida que mais e mais informação é adquirida a partir do campo da Genômica, a questão ganhará, sem dúvida, maior atenção. Um exemplo de um livro detalhando a evidência convincente de DNA para a evolução é The Making of the Fittest: DNA and the Ultimate Forensic Record of Evolution (A criação dos mais aptos: DNA e o registro forense final da evolução). Uma descrição do editor inclui os seguintes comentários:
A evidência de DNA não apenas soluciona crimes — nas mãos de Sean Carroll, isso vai acabar com as Guerras da Evolução.
O DNA é o material genético que nos define como indivíduos. Ao longo das duas últimas décadas, surgiu como uma ferramenta poderosa para solucionar crimes e determinar culpa e inocência. Mas, muito recentemente, um novo aspecto importante do DNA foi revelado – contém um registro detalhado da evolução. Isto é, o DNA é uma crônica viva de como as criaturas maravilhosas que habitam nosso planeta adaptaram-se aos seus muitos ambientes, das águas congelantes da Antártida à copa exuberante da floresta tropical.
Nas páginas desta narrativa de leitura altamente agradável, Sean Carroll guia o leitor em geral em uma turnê de um registro maciço de DNA de três bilhões de anos de evolução para ver como os aptos são feitos. E que turnê de abrir os olhos – uma com genes imortais, genes fósseis e genes que carregam as cicatrizes de batalhas passadas com doenças horríveis. Este livro encerra o caso para a evolução, para além de qualquer dúvida razoável.
Parece que será apenas uma questão de tempo antes dos argumentos anti-evolucionistas baseados no registro fóssil e na “Explosão Cambriana” serem “superados” por mais dados definitivos da pesquisa de DNA. É necessário que os apologistas cristãos modernos aprendam as lições da história para que sejam parte da solução em vez de ser parte do problema. Ganhar um entendimento mais profundo destas questões será fundamental se queremos manter a credibilidade necessária para ajudar outros a ver a harmonia entre a revelação natural e a especial de Deus.
John Lang
¹ : http://download.audible.com/product_related_docs/BK_SANS_000715.pdf
²: http://pt.wikipedia.org/wiki/Origem_comum
³: Francis Collins, A Linguagem de Deus, (Nova Iorque: Free Press, 2006), 124-141 (vários extratos)
*: Fazale Rana & Hugh Ross, Quem foi Adão?, (Colorado Springs: NavPress, 2005)
**: http://www.asa3.org/ASA/topics/Book%20Reviews2005-/12-05.html
***: Fazale Rana & Hugh Ross, Quem foi Adão?, (Colorado Springs: NavPress, 2005), 243
Fonte: http://www.douglasjacoby.com/wp-content/uploads/HGP%20&%20Apologetics.pdf
Tradução: Mariana Maia